Nikita V. Zelenkov, 2021
New Bird Taxa (Aves: Galliformes, Gruiformes) from the Early Eocene of Mongolia
Paleontological Journal 55: 438-446
Free pdf:
(PDF) New Bird Taxa (Aves: Galliformes, Gruiformes) from the Early Eocene of Mongolia
SYSTEMATIC PALEONTOLOGY
Order
Galliformes
Galliformes Incertae Familiae
Genus
Bumbanortyx Zelenkov, gen. nov.
Etymology. From Bumban, the lower Eocene member in the Tsagaan-Khushuu locality, and the Latin
Ortyx (quail), feminine.
Type species. Bumbanortyx transitoria sp. nov.
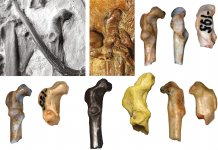
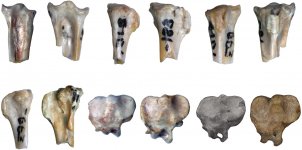
Diagnosis. The cotyla scapularis of the coracoid is large, vaguely oval, mostly dorsally oriented; the medial margin of the shaft at the level of the facies articularis humeralis is oriented subparallel to the long axis of the coracoid (somewhat inclined medially) and forms a shallow notch near the processus acrocoracoideus; the processus acrocoracoideus greatly projects cranially relative to the apex of the facies articularis
humeralis and medially relative to the medial margin of the shaft (owing to the medial displacement of the impressio bicipitalis); the cranial part (apex) of the impressio lig. acrocoracohumeralis is oriented dorsocranially and is visible in dorsal view, the caudal part of the impressio is oriented cranially; the facies articularis clavicularis is craniocaudally expanded; the impressio bicipitalis markedly projects ventrally relative to the adjacent ventral surface of the shaft; the facies articularis humeralis is short (its length roughly twice its width), wide and has a blunt cranial end; the labrum glenoidale greatly projects ventrally and is convex; the processus procoracoideus is moderately well developed (its width roughly corresponds to half the width of the cotyla scapularis).
The humerus has a craniocaudally narrow caput humeri that does not form a ventral expansion; the bony bridge connecting the ventral part of the caput and the tuberculum ventrale is absent; the tuberculum dorsale is elongated; the fossa tricipitalis dorsalis is present, though not deep, the caput humeri overhangs it slightly; the fossa tricipitalis is not pneumatized.
The tarsometatarsus has weakly developed fossae parahypotarsales (the lateral one is barely noticeable), the sulcus extensorius is fairly deep, delimited by a thin medial margin and a thickened lateral margin, proximally passing into the deep fossa infracotylaris dorsalis; the hypotarsus is proximodistally short, dorsoplantarly low, with a width that is 2/3 the width of the proximal end; the canal for the m. flexor digirotum longus is located near the plantar margin of the hypotarsus (not dorsally embedded); the grooves for the tendons of the flexor muscles of the II digit and the m. flexor hallucis longus are oriented plantarly, the groove for the m. flexor hallucis longus is located somewhat dorsal to the former. In dorsal view, the lateral margin of the shaft is oriented subparallel to the long axis of the bone, whereas the medial margin forms an inflection at the level of the fossa infracotylaris dorsalis.
Species composition. Type species.
Comparison. The coracoid of
Bumbanortyx resembles
Quercymegapodius Mourer-Chauviré, 1992 in general outline, but differs from it in having a fairly large processus procoracoideus (completely reduced in
Quercymegapodius: Mourer-Chauviré, 1992), a shallow but distinct notch on the medial margin of the bone shaft immediately next to the processus acrocoracoideus (Fig. 1, inc), as well as a shaft that does not expand medially in the cranial direction (in
Bumbanortyx, the medial margin of the shaft is subparallel to the long
axis of the bone at the level of the facies articularis humeralis).
Bumbanortyx differs from genera
Gallinuloides Eastman, 1900 and
Paraortygoides Mayr, 2000 (Gallinuloididae) in having a more rounded cotyla scapularis (distinctly oval in
Gallinuloides and
Paraortygoides: Mayr, 2000; Mayr and Weidig, 2004), a thinner and more transversely oriented crista acrocoracoidea (thicker, bulge-shaped and more longitudinally oriented in
Gallinuloides and
Paraortygoides; Fig. 1, ca) and a somewhat shortened extremitas omalis (in
Gallinuloides and
Paraortygoides, the part of the coracoid cranial to the cotyla scapularis is elongated; Figs. 1a, 1b).
Bumbanortyx differs from Paraortygidae
in the rounder cotyla scapularis (in Paraortygidae, including
Xorazmortyx Zelenkov et Panteleyev, 2019, the long axis of the cotyla is oriented obliquely relative to the long axis of the bone: Mourer-Chauviré, 1992; Zelenkov and Panteleyev, 2019; Stidham et al., 2020).
Bumbanortyx further differs from the type genus
Paraortyx Gaillard, 1908 in having an impressio lig. acrocoracohumeralis that is oriented dorsally in dorsal view and the processus acrocoracoideus that entirely projects cranially in dorsal view.
Bumbanortyx differs from
Xorazmortyx and
Scopelortyx Mourer-Chauviré, Pickford et Senut, 2015, which have a coracoid that is morphologically different from
Paraortyx, in having a
medially expanded processus acrocoracoideus in dorsal view (see Mourer-Chauviré et al., 2015; Zelenkov and Panteleyev, 2019).
The humerus of
Bumbanortyx differs from all other galliform families in the narrow (not craniocaudally expanded) caput humeri (its condition is unknown in
Gallinuloides). It differs from the early Eocene
Paraortygoides (Gallinuloididae) in the shallow fossa tricipitalis dorsalis.
The tarsometatarsus of
Bumbanortyx differs from
Paraortygoides in the more centrally placed hypotarsus (in
Paraortygoides it is laterally displaced: Mayr, 2016, text-fig. 2E); it differs from members of the families Quercymegapodiidae and Paraortygidae in the dorsoplantarly low hypotarsus. Differs from
Argillipes aurorum Harrison et Walker, 1977 in the mediolaterally wider hypotarsus due to the more plantar orientation of the sulcus for m. flexor hallucis longus (in
Argillipes aurorum the hypotarsus is narrower because the sulcus for m. flexor hallucis longus is generally oriented laterally; Harrison and Walker, 1977; Mayr and Smith, 2019). Also, in
Argillipes aurorum, the medial margin of the tarsometatarsus does not form an expansion at the transition with the proximal end, whereas in
Bumbanortyx a distinct inflection is present at the level of the fossa infracotylaris dorsalis. Differs from
Percolinus Harrison et Walker, 1977 in having a dorsoplantarly low hypotarsus and an inflection in the medial margin at the transition to the proximal end.
Remarks. Bumbanortyx represents a stem galliform, because it has a deeply excavated rounded cotyla scapularis, as in the fossil families Paraortygidae, Gallinuloididae and Quercymegapodiidae (Mourer-
Chauviré, 1992; Mayr, 2000). In general outlines and morphology
Bumbanortyx is closest to
Quercymegapodius, with which it shares a large, rounded and strictly dorsally located cotyla scapularis (in Paraortygidae and Gallinuloididae, the cotyla scapularis is oval and
somewhat displaced laterally), a strongly cranially projecting processus acrocoracoideus, as well as a short, wide and strongly ventrally projecting (forming a high labrum glenoidale) facies articularis humeralis (see Mourer-Chauviré, 1992; Alvarenga, 1995). In
Quercymegapodius, the processus procoracoideus is completely reduced, but it is present in
Ameripodius silvasantosi Alvarenga, 1995 from the Oligocene of Brazil, which also shares with
Bumbanortyx a straight shaft in the cranial part of the bone (see Alvarenga, 1995). The absence of processus procoracoideus was previously viewed as a typical trait of Quercymegapodiidae (Mourer-Chauviré, 1992), but the presence of a pronounced processus procoracoideus in
Ameripodius
silvasantosi and, to lesser extent, in
A. alexis Mourer-Chauviré, 2000 shows that this trait is variable in Quercymegapodiidae.
The presence of a shallow though distinct notch on the medial margin of the coracoid shaft near the processus acrocoracoideus and a relatively large processus procoracoideus suggest an affinity between
Bumbanortyx and Gallinuloididae, though structurally
Bumbanortyx
is similar to Quercymegapodiidae. The coracoid of
Bumbanortyx therefore shows a mosaic of morphological characters of Gallinuloididae and Quercymegapodiidae and can be considered as the
intermediate link between these two families. Because Gallinuloididae are known from the early Eocene, while Quercymegapodiidae are only known from the late Eocene (Mourer-Chauviré, 1992; Mayr, 2009), the morphological characters shared by
Bumbanortyx and Gallinuloididae can be plesiomorphic, whereas those shared by
Bumbanortyx and Quercymegapodiidae are obviously advanced. Nevertheless, the intermediate morphology of Bumbanortyx shows that the morphological type of the coracoid in Quercymegapodiidae evolved from that of the Gallinuloididae.
Another coracoid from the Bumban Member (IMG 100/1371) was described as a probable member of the family Quercymegapodiidae (Hood et al., 2019). IMG 100/1371 generally has similar morphology to the one described in this article, but differs in its larger size and the absence of a notch on the medial margin of the shaft, so that it may represent a separate species.
The most characteristic trait of
Bumbanortyx is its narrow caput humeri (lacking a ventral expansion), which is not typical for any other known members of Galliformes. This character was previously noted for the unnamed galliform bird from the Bumban Member (Hwang et al., 2010), which I assign here to
Bumbanortyx. The presence of the same morphology of caput humeri was recently noted for a small galliform from the lower Eocene of Belgium, tentatively assigned to
Argillipes (Mayr and Smith, 2019). The presence of this character in several early Eocene galliforms may indicate that they form a separate familylevel group. It should also be noted that although the morphology of the caput humeri is unknown for
Gallinuloides,
Bumbanortyx partly resembles it in the morphology of the coracoid, and thus it cannot be ruled out that
Bumbanortyx belongs to Gallinuloididae. On the other hand, the general structural similarity with the coracoid of the younger genus
Quercymegapodius may be apomorphic, in which case
Bumbanortyx may turn out to be a primitive representative of Quercymegapodiidae.
Tarsometatarsi assigned to
Bumbanortyx transitoria, resemble those of
Argillipes aurorum in general proportions but differ in the shape of the hypotarsus and the presence of an inflection on the medial margin of the bone at the transition to the proximal end (see Comparison above). A small galliform is known from the early Eocene of Belgium that resembles
Argillipes aurorum in the morphology of the tarsometatarsus, and also has a narrow caput humeri (Mayr and Smith, 2019). The Belgian bird also has the characteristic inflection typical for
Bumbanortyx, but a different morphology of the hypotarsus. The unnamed Belgian taxon and
Argillipes aurorum therefore are likely both
phylogenetically close to
Bumbanortyx transitoria. Differences from
Paraortygoides in the morphology of the tarsometatarsus preclude affinities of
Bumbanortyx with Gallinuloididae.
“Argillipes” paralectoris Harrison et Walker, 1977 likely represents a separate genus, and its assignment to Galliformes requires confirmation (Mayr and Smith, 2019).
In the general organization of the proximal end of the tarsometatarsus and in particular of the hypotarsus,
Bumbanortyx resembles the extant Cracidae, from which it differs in the plantar position and the small diameter of the canal for the m. flexor digitorum longus, as well as the almost undeveloped fossae parahypotarsales. The latter character reflects poorly developed short muscles of the foot, and, therefore, a different locomotor specialization of
Bumbanortyx compared with the extant Cracidae.
Bumbanortyx transitoria Zelenkov, sp. nov.
Etymology. From the Latin transitorius (transitional).
Holotype. PIN, no. 3104/265, cranial fragment of left coracoid; Mongolia, Tsagaan-Khushuu locality; lower Eocene, Bumban Member. Collected by JSMPE in 1980.
Description (Figs. 1c, 1d, 1f, 1j; 2a, 2c, 2e, 2g, 2i; 3a, 3b). Same as the diagnosis for the genus (which also serves as the diagnosis of the new species).
Measurements (in mm). Coracoid: length from the cranial apex of the bone to the caudal margin of the cotyla scapularis 5.6; dorsoventral height at the level of the cotyla scapularis 2.2. Humerus: width of the proximal end 7.8; craniocaudal height of the caput humeri 2.5. Tarsometatarsus: width of the proximal end 4.4.
Comparison. The genus
Bumbanortyx has one species.
Remarks. Based on similar relative size I assign the humerus and the tarsometatarsus to
B. transitoria. The dorsoventral width of the caput humeri in PIN, no. 3104/663 and the length of the glenoid part of the coracoid (from the apex of the facies articularis humeralis to the tip of the processus procoracoideus) in the holotype correspond to small specimens of the extant Coturnix coturnix. The second galliform bird from Tsagaan-Khushuu (Bumbanipodius magnus gen. et sp. nov.) is distinctly larger.
Material. In addition to the holotype, includes the following specimens from the type locality: PIN, no. 3104/128, proximal fragment of left humerus; PIN, no. 3104/663, proximal fragment of left tarsometatarsus.
Fred