Systematic paleontology
Aves Linnaeus, 1758.
Galliformes Temminck, 1820.
Gen. et sp. indet.
Referred specimen
SMF Av 666: partial cranium
Locality and horizon
Beach terrace approximately 320 m northwest of the mouth of Jansen Creek, south shore of the Strait of Juan de Fuca, Clallam County, Washington State, USA (48.32665° N, 124.47338° W); Jansen Creek Member of the Makah Formation, latest Eocene or earliest Oligocene; collected by Terry Iversen on 16th June 2017.
Measurements (in mm)
Length as preserved, 37.5; estimated total length of complete skull (assuming a beak with galliform-like proportions), 55‒60; width across processus postorbitales, 25.7; width of nasofrontal hinge, 13.3.
Description and comparisons
The specimen consists of a cranium lacking the palatal bones as well as the rostrum, broken away at the nasofrontal hinge. As indicated by the completely co-ossified sutura frontoparietalis, it is from an adult bird (fissures along the dorsolateral surfaces represent cracks owing to the fact that the fossil was fractured in the discovery process). Because skulls of other Paleogene galliforms, that is, the gallinuloidid taxa Paraortygoides and Gallinuloides and the phasianoidean taxon Palaeortyx, are too poorly preserved for detailed comparisons, the following description is mainly based on comparisons with crown group Galliformes.
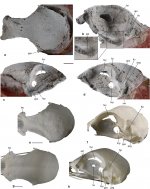
The fossil agrees well with the skulls of some crown group Galliformes in proportions and morphological features (Fig. 1). No traces of an ectethmoid can be discerned, but the corresponding area is not well preserved (the ectethmoid is well developed in the Megapodiidae but reduced in the Cracidae and Phasianidae). Likewise, there are no well-defined articular facets for the lacrimals (in the Cracidae, the caudal portion of the lacrimal articulates with the frontal and is attached to the orbital rim; Fig. 1g).
Unlike in the Megapodiidae, the fonticulus orbitocranialis is well developed; in contrast to the Cracidae (Fig. 1h), there is no laterally open sulcus olfactorius. On the left side of the specimen, a kidney-shaped foramen is located caudal of the fonticulus orbitocranialis (Figs. 1d, 2g); this foramen is not present on the right side of the fossil. A fonticulus interorbitalis is absent. The large foramen opticum is similar in size and position to the foramen of crown group Galliformes. The interorbital section formed by the frontal bones is mediolaterally narrow and measures only about one third of the maximum width of the skull. The interorbital section is equally narrow in many taxa of the Phasianidae (Fig. 1e), whereas it is wider in the Megapodiidae, Cracidae (Fig. 1g), and Numididae. The cranium has a sub-cylindrical shape and its dorsal surface is not as vaulted as it is in most crown group Galliformes. An equally flat dorsal surface of the cranium is found in the phasianid taxa Ammoperdix and Alectoris. Cross sectional images generated by micro-CT scans show that the bone walls of the dorsal portion of the neurocranium are very thick in the area of the parietals, as they are in at least some crown group Galliformes (only a cross section of the skull of Gallus gallus was available to us), whereas the skull roof of the fossil has thinner bone walls than G. gallus in the area of the frontals (Fig. 3a‒d).
As in other galliforms, the processus postorbitalis is a laterally protruding, subtriangular projection rather than a rod-shaped process. In the fossil, the tip of the right processus postorbitalis is broken, whereas the left one is complete.
The lateral margin of the processus postorbitalis forms a ridge, as it does in many crown group Galliformes. Overall, the morphology of the temporal region of the fossil (Fig. 4a) is most similar to that of extant Arborophila (Fig. 4g), which is one of the earliest diverging taxa of the
Phasianidae (Wang et al. 2013). The processus zygomaticus is a dorsoventrally oriented, flange-like embossment, and there is no indication of an ossified aponeurosis zygomatica (Fig. 4f, i), which characterizes many crown group Galliformes (Zusi and Livezey 2000; Elzanowski and Mayr 2018). An ossified aponeurosis zygomatica evolved multiple times within crown group Galliformes (see discussion) and is absent in the early Eocene stem group galliform Gallinuloides (Mayr and Weidig 2004) and the Oligocene Palaeortyx (Mayr et al. 2006), which is an early diverging representative of the Phasianoidea. A short processus suprameaticus is visible on the left side of the fossil, immediately caudal of the cotyla quadratica squamosi (of which only the lateral rim is exposed). Unfortunately, the tip of this process is broken, so we cannot determine whether it contributed to an osseous
bridge delimiting the cranial margin of the tympanic cavity, as it does in many crown group Odontophoridae (except Ptilopachus; Fig. 4e) and Phasianidae (including Arborophila; Fig. 4g).
As in most extant Galliformes, the fossa temporalis is poorly developed. Furthermore, as in crown group Galliformes, there is a small vascular foramen in this fossa, between the processus postorbitalis and the processus zygomaticus (Fig. 4; see discussion). This foramen is of similar size and position to the foramen in crown group Galliformes and is macroscopically visible (Fig. 4a). Owing to the fact that it is infilled by sediment of similar density to the bone, the foramen is not well delimited in the surface model created by micro-CT data (Fig. 2h), even though cross sectional images of this area show a canal that passes into the brain cavity (Fig. 2j). Unlike in extant galliform birds, the fossil exhibits a marked, pit-like depression rostral to this foramen (Figs. 2h, 4a).
The rostrum parasphenoidale exhibits rostrocaudally elongate and sessile processus basipterygoidei (Figs. 1b, 2f), a derived character of galloanserine birds. In the fossil, the articular facets of these processes are poorly developed, which is especially true for the right one that appears to be eroded. Other features of the basicranium, as well as details of the otic region, are not preserved in the fossil. An elongate element situated next to the left otic region (Fig. 1d) cannot be identified. This structure may be mistaken for an ossified aponeurosis zygomatica, but in contrast to the latter, it is not flattened and sheet-like but tubular. The apparent bifurcation of this element and its clear separation from the processus zygomaticus furthermore preclude its
identification as an ossified aponeurosis zygomatica. Judging from its slightly darker color, it may not be bone but a small piece of wood; fossilized wood is occasionally found together with avian remains in the Makah Formation. The virtual cranial endocast (Fig. 5a, b) exhibits a morphology similar to that of extant Megapodiidae and Phasianidae
(Fig. 5c‒f; Handley and Worthy 2021, see also Early et al. 2020). As in the latter, the shapes of the two telencephalic hemispheres give the rostral portion of the endocast a heart-shaped outline in dorsal and ventral view. In addition, Fig. 2 a‒g Micro-CT scans of the fossil cranium (SMF Av 666) in a left lateral, b right lateral, c cranial, d dorsal, e ventral, and f rostrolateral view; g shows a detail of the framed area in f. h‒j Detail of temporal region with transverse section on the level of the foramen temporale venosum; in i, the plane of the section is shown. dpr pit-like depression rostral to foramen temporale venosum, for kidneyshaped foramen caudal of fonticulus orbitocranialis, ftv foramen temporale venosum, pbt processus basipterygoideus, ppo processus postorbitalis, pzg processus zygomaticus. Scale bar equals 5 mm in a‒f
and 1 mm in g‒j. [Color online] as in crown group Galliformes, the wulsts are weakly developed. The mesencephalon (optic lobe) and cerebellum are likewise of similar shape and size to the corresponding brain parts of crown group Galliformes. Whereas there was a clear differentiation between the bones and the rostral portion of the endocast (telencephalic hemispheres) in the micro-CT scans, no unambiguous distinction between bone and surrounding matrix was possible in the area of the flocculus and our reconstruction of the shape of the latter structure is tentative. The bulbus olfactorius is not completely preserved.
Fred
Fig. 1 a‒d Cranium of a galliform bird (SMF Av 666) from the latest
Eocene or early Oligocene Jansen Creek Member of the Makah Formation, Washington State, USA, in a dorsal, b rostrolateral, c right lateral, and d left lateral view; the caudal portion of the specimen is attached to synthetic resin (red color). e, f Cranium of the extant Arborophila brunneopectus (Phasianidae; SMF 14588) in e dorsal
and f left lateral view. g, h Cranium of the extant Ortalis canicollis
(Cracidae; SMF 14247) in g dorsal and h left lateral view. dpr pitlike
depression rostral to foramen temporale venosum, fio fonticulus
interorbitalis, foc fonticulus orbitocranialis, fop foramen opticum,
for kidney-shaped foramen caudal of fonticulus orbitocranialis, ftp
fossa temporalis, ftv foramen temporale venosum, lac lacrimal, pbt
processus basipterygoideus, ppo processus postorbitalis, pzg processus
zygomaticus, sof sulcus olfactorius, uie unidentified element. The
fossil specimen was coated with ammonium chloride. The scale bars
equal 5 mm. [Color online]