Acanthis
Well-known member
Hahaha oh well!Nomen nudum and , probably, nomen oblitum. To o-bli-viooooon !




Hahaha oh well!Nomen nudum and , probably, nomen oblitum. To o-bli-viooooon !

Chlorochroa Stål, 1872 is protected, in theory. But, in my knowledge, Chlorochroa Sclater, 1862, was never published after 1899.Hahaha oh well!
Chlorochroa Stål, 1872 is protected, in theory. But, in my knowledge, Chlorochroa Sclater, 1862, was never published after 1899.
Welcome Eurocephalidae
But it will be, and does not constitute published work within the meaning of the Code as per ICZN 9.9.

I hadn't even seen it !!The recommendations in the study:
"The levels of divergence between the three major populations of M. lugubris are well above those at which ornithologists would normally assign species rank. Consequently, we tentatively propose that these three populations should be elevated to species rank, M. lugubris (Schlegel, 1871) in the Vogelkop region, Melampitta rostrata (Ogilvie-Grant, 1913) in the western central range and Melampitta longicauda (Mayr & Gilliard, 1952) in the eastern central range."

It would be good to have Neosittidae with Neositta in it, that's for sure - but only c. 5 million years of separation. Extra support for paraphyly of D. chrysoptera more noteworthy for me - and ancient divergence within Alaeadryas, Cinclosoma ajax, Pachycephala soror & Melanorectes. Interesting paper, thanks.
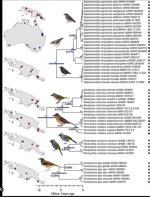
I finally found the article which, for me, can justify the separation of Daphoenositta into two genera Daphoenositta and Neositta (figure below)
For me, the 3 Daphoenositta look very distinct in plumage despite the small divergence time (I learned that divergence time doesn't actually mean anything) and the two main clades are reciprocally monophyletic. Finally, apart from genetic distance, there are no arguments in favor of a single genus just as there are no arguments in favor of two genera. In this case, it is purely arbitrary and the monophyly is not disturbed anyway.Not really sure how you interpret that figure to suggest they are different genera? What am I missing?
They're only mtDNA data though: I'd prefer more genetic back-up before splitting up Varied Sittella. On plumage, these clusters still do not make a lot of sense!It would be good to have Neosittidae with Neositta in it, that's for sure - but only c. 5 million years of separation. Extra support for paraphyly of D. chrysoptera more noteworthy for me - and ancient divergence within Alaeadryas, Cinclosoma ajax, Pachycephala soror & Melanorectes. Interesting paper, thanks.
Agree we need more data before anything official happens but I enjoy the speculationThey're only mtDNA data though: I'd prefer more genetic back-up before splitting up Varied Sittella. On plumage, these clusters still do not make a lot of sense!
")
Bonaccorso & al (2009) have suggested to raised Cyanolyca viridicyanus jolyaea to species but they don't proposed a name. Has an English name been proposed for this taxon that I could translate?
Simply. I was expecting a more elaborate name 😂😂Oops.v.13:pt.7 (1934) - Catalogue of birds of the Americas and the adjacent islands in Field Museum of Natural History - Biodiversity Heritage Library
The Biodiversity Heritage Library works collaboratively to make biodiversity literature openly available to the world as part of a global biodiversity community.www.biodiversitylibrary.org