


-
Welcome to BirdForum, the internet's largest birding community with thousands of members from all over the world. The forums are dedicated to wild birds, birding, binoculars and equipment and all that goes with it.
Please register for an account to take part in the discussions in the forum, post your pictures in the gallery and more.

You are using an out of date browser. It may not display this or other websites correctly.
You should upgrade or use an alternative browser.
You should upgrade or use an alternative browser.
Cuckoos of the World (1 Viewer)
- Thread starter Richard Klim
- Start date
More options
Who Replied?andrew147
Well-known member
Crested, Blue and Verraux's all are arboreal and Serisomus Swainson, 1837 (type species = C. cristata) seems good for those. Not sure if Delalande's was arboreal but in 1854 Bonaparte put it in Serisomus. See page 5 of https://books.google.com/books?id=TfFhAAAAcAAJ&pg=PP8&dq=huskiricmis#v=onepage&q=Serisomus&f=false
Thanks for replies. I will continue to use Serisomus.
As for Delalande's Coua, it is included in the comprehensive trees of Payne et al (2005) and placed as sister to Coua serriana, this pair closest in turn to C. gigas - so, embedded well within the terrestrial group.
Last edited:
Jim LeNomenclatoriste
Je suis un mignon petit Traquet rubicole

Glaucococcyx. Cabanis in Mus. Hein. IV. 1. 1863 = Glaucomyias, Cabanis in Mus. Heiu. 1. 1850.
I'm not sure that Glaucococcyx and Glaucomyias are synonyms. Glaucomyias is, IMHO, a synonym of Eumyias (Muscicapidae)?
mb1848
Well-known member
‘As for Delalande's Coua, it is included in the comprehensive trees of Payne et al (2005) and placed as sister to Coua serriana, this pair closest in turn to C. gigas - so, embedded well within the terrestrial group”
After a little research I think that Delalande’s Coua is an aberrant arboreal coua. The arboreal ones eat insects and land mollusks the ground ones eat insects and seeds. Delalande’s eat land mollusks; their native name meaning shell cracker.
https://digital.soas.ac.uk/LOAC000068/00015/92j?search=coua .
This source considers C. serriana and reynaudii as arboreal???
C. delandei was described in Coccycus note the c instead of the z by Temminck. Apparently a misspelling but perhaps Temminck meant to create a new genus. Our dear friend Hans E. Wolters had his own ideas. He placed C. reynaudii and serriana (both Pucheran 1845) in an unnamed subgenus. Wolters put C. caerulea in subgenus Glaucococcyx Cabanis & Heine, this makes sense since although arboreal and land mollusk eating their legs are the length of the terrestrial couas. Wolters put C. verreaauxi and cristata in a subgenus Serisomus.
After a little research I think that Delalande’s Coua is an aberrant arboreal coua. The arboreal ones eat insects and land mollusks the ground ones eat insects and seeds. Delalande’s eat land mollusks; their native name meaning shell cracker.
https://digital.soas.ac.uk/LOAC000068/00015/92j?search=coua .
This source considers C. serriana and reynaudii as arboreal???
C. delandei was described in Coccycus note the c instead of the z by Temminck. Apparently a misspelling but perhaps Temminck meant to create a new genus. Our dear friend Hans E. Wolters had his own ideas. He placed C. reynaudii and serriana (both Pucheran 1845) in an unnamed subgenus. Wolters put C. caerulea in subgenus Glaucococcyx Cabanis & Heine, this makes sense since although arboreal and land mollusk eating their legs are the length of the terrestrial couas. Wolters put C. verreaauxi and cristata in a subgenus Serisomus.
andrew147
Well-known member
After a little research I think that Delalande’s Coua is an aberrant arboreal coua.
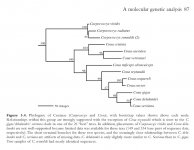
I've attached the tree from PAYNE, R., SORENSON, M. & KLITZ K. (2005) The Cuckoos: Cuculidae; Oxford University Press.
I can't say if it's definitive - certainly Verreaux's being closer to Blue than to Crested is unexpected... and note what it says in the legend about C. delalandei with regard to C. serriana/C. gigas. However, the division into a 'mostly-terrestrial' group and a 'mostly-arboreal' group based on genetics, also seems reasonable to me in terms of morphology, taking into account general proportions, overall colouration, crestedness - and also feeding behaviour.
Based on info in Erritzøe et al (2012), Payne et al, and also HBW, all extant couas have rather generalised diets (reptiles, insects, fruit, seeds); only C. delalandei was specialised, eating "large forest snails broken on stone anvils". Furthermore, Erritzøe et al and Payne et al both state that C. delalandei was "terrestrial" - there doesn't seem much to indicate that it was actually related to the arboreal spp.
Attachments
Last edited:
Peter Kovalik
Well-known member

BirdLife Taxonomy: Dusky Long-tailed Cuckoo (Cercococcyx mechowi) is being split: assessment of newly recognised taxa.
Following a taxonomic reassessment, Dusky Long-tailed Cuckoo (Cercococcyx mechowi) has been split into Dusky Long-tailed Cuckoo (C. mechowi), and Whistling Long-tailed Cuckoo (C. lemaireae) (see Boesman and Collar, 2019). The newly-split Dusky Long-tailed Cuckoo has a range extending from central Cameroon east to Uganda. Whistling Long-tailed Cuckoo ranges from Sierra Leone to western Cameroon (Boesman and Collar, 2019).
The exact habitat requirements for the newly-split Whistling Long-tailed Cuckoo have not been investigated, but it is very likely to require lowland and montane forests, and have the same high forest dependency as Dusky Long-tailed Cuckoo (del Hoyo et al., 2002). This high forest dependency may render these species vulnerable to forest loss. The population size of the pre-split species is unknown, but it is described as relatively uncommon but not rare (del Hoyo et al., 2002).
The pre-split species was previously listed as Least Concern (BirdLife International 2020). However, following the taxonomic split, new range sizes suggest that both species warrant a thorough reassessment. We have therefore reassessed both species against each criterion here.
Following a taxonomic reassessment, Dusky Long-tailed Cuckoo (Cercococcyx mechowi) has been split into Dusky Long-tailed Cuckoo (C. mechowi), and Whistling Long-tailed Cuckoo (C. lemaireae) (see Boesman and Collar, 2019). The newly-split Dusky Long-tailed Cuckoo has a range extending from central Cameroon east to Uganda. Whistling Long-tailed Cuckoo ranges from Sierra Leone to western Cameroon (Boesman and Collar, 2019).
The exact habitat requirements for the newly-split Whistling Long-tailed Cuckoo have not been investigated, but it is very likely to require lowland and montane forests, and have the same high forest dependency as Dusky Long-tailed Cuckoo (del Hoyo et al., 2002). This high forest dependency may render these species vulnerable to forest loss. The population size of the pre-split species is unknown, but it is described as relatively uncommon but not rare (del Hoyo et al., 2002).
The pre-split species was previously listed as Least Concern (BirdLife International 2020). However, following the taxonomic split, new range sizes suggest that both species warrant a thorough reassessment. We have therefore reassessed both species against each criterion here.
Jim LeNomenclatoriste
Je suis un mignon petit Traquet rubicole

Jim LeNomenclatoriste
Je suis un mignon petit Traquet rubicole
Cacomantis addendus Rothschild & Hartert, 1901 – FR: Coucou des Salomon/ENG: Solomon Cuckoo
Cacomantis blandus Rothschild & Hartert, 1914 – FR: Coucou de l'Amirauté/ENG: Admiralty Cuckoo
Cacomantis virescens (Brüggeman, 1876) – FR: Coucou de Brüggeman/ENG: ?Brüggeman's Cuckoo
Last edited:
sicklebill
well-known Cretaceous relic

They split 6 species out as follows:
Integration of the resulting data set distinguished six species-level taxa within the complex: sepulcralis in the Philippines and Sundaland east to central Nusa Tenggara, virescens in Sulawesi and the Sula Archipelago, aeruginosus in the Moluccas, variolosusin east Nusa Tenggara, the Moluccas, north and east Australia, New Guinea and Bismarck Archipelago, blandus in the Admiralty Islands, and addendusin the Solomon Islands. Our review of infra-specific differentiation among species leads us to distinguish 13 subspecies.
Now for someone to tackle Fan-tailed Cuckoo.........
Integration of the resulting data set distinguished six species-level taxa within the complex: sepulcralis in the Philippines and Sundaland east to central Nusa Tenggara, virescens in Sulawesi and the Sula Archipelago, aeruginosus in the Moluccas, variolosusin east Nusa Tenggara, the Moluccas, north and east Australia, New Guinea and Bismarck Archipelago, blandus in the Admiralty Islands, and addendusin the Solomon Islands. Our review of infra-specific differentiation among species leads us to distinguish 13 subspecies.
Now for someone to tackle Fan-tailed Cuckoo.........
l_raty
laurent raty
They split 6 species out as follows:
Cacomantis sepulcralis - Plazi TreatmentBank
Cacomantis virescens - Plazi TreatmentBank
Cacomantis aeruginosus Salvadori 1878 - Plazi TreatmentBank
Cacomantis variolosus - Plazi TreatmentBank
Cacomantis blandus Rothschild & Hartert 1914 - Plazi TreatmentBank
Cacomantis addendus Rothschild & Hartert 1901 - Plazi TreatmentBank
(That being said, if someone has the actual pdf, I'd be interested to see it.
") )
)sicklebill
well-known Cretaceous relic
So we have two splits in Brush Cuckoo C. variolosus- Admiralty Brush Cuckoo C. blandus and Solomons Brush Cuckoo C. addendus. Two potential lifers anyway out of that anyway in these tick deficient times.
C. sepulcalis is split as Rusty-breasted Cuckoo by IOC with virescens being a split from that species; C. aeruginosus is an IOC species already as Moluccan Cuckoo.
C. sepulcalis is split as Rusty-breasted Cuckoo by IOC with virescens being a split from that species; C. aeruginosus is an IOC species already as Moluccan Cuckoo.
Peter Kovalik
Well-known member
Sánchez-González, L.A., Cayetano, H., Prieto-Torres, D.A., Rojas-Soto, O.R., and Navarro-Sigüenza, A.G. (2002) The role of ecological and geographical drivers of lineage diversification in the Squirrel cuckoo Piaya cayana in Mexico: a mitochondrial DNA perspective. Journal of Ornithology. Published online 15 August 2022.
https://doi.org/10.1007/s10336-022-02008-w
Abstract
The Squirrel Cuckoo (Piaya cayana) is a widely distributed neotropical species with 14 recognized subspecies. Two parapatric subspecies are distributed in Mexico. P. c. mexicana is endemic to the seasonally dry tropical forests of western Mexico, and P. c. thermophila is found in humid forests from eastern Mexico to western Colombia. The 2 taxa have a small area of overlap on the Isthmus of Tehuantepec, but there is pronounced phenotypic differentiation, and very few reported intermediate individuals between these forms, leading to debate about the taxonomic status of the Mesoamerican taxa. Based on two mitochondrial genes, we used phylogenetic, phylogeographic, morphological, and ecological modelling analyses to describe the evolutionary relationships and paleo-distributional patterns of P. cayana in Mexico. Divergence time estimates suggest that the split between P. c. mexicana and P. c. thermophila occurred during the Pleistocene, about 1.24 Mya. Morphometrics of Mexican subspecies indicate that tail length and the white tail tips are significantly longer in P. c. mexicana, while P. c. thermophila has a longer and wider bill. Ecological niche analyses indicated that niche similarity between both lineages was lower than expected by chance, while low values on cross-prediction tests suggested that the two lineages have inhabited different environmental spaces since at least the Late Pleistocene. The ecological niche difference may also be associated with a steep humidity gradient, suggesting a significant contemporary environmental barrier. Overall, our results indicate that P. c. mexicana and P. c. thermophila have divergent evolutionary histories; therefore, the current taxonomic status of the Piaya populations in Mexico reflects neither their evolutionary relationships nor their apparent divergence. Our results suggest a major split in the polytypic P. cayana coinciding with the Andes, suggesting that the western endemic P. c. mexicana and P. c. thermophila are best treated as separate species-level taxa.
https://doi.org/10.1007/s10336-022-02008-w
Abstract
The Squirrel Cuckoo (Piaya cayana) is a widely distributed neotropical species with 14 recognized subspecies. Two parapatric subspecies are distributed in Mexico. P. c. mexicana is endemic to the seasonally dry tropical forests of western Mexico, and P. c. thermophila is found in humid forests from eastern Mexico to western Colombia. The 2 taxa have a small area of overlap on the Isthmus of Tehuantepec, but there is pronounced phenotypic differentiation, and very few reported intermediate individuals between these forms, leading to debate about the taxonomic status of the Mesoamerican taxa. Based on two mitochondrial genes, we used phylogenetic, phylogeographic, morphological, and ecological modelling analyses to describe the evolutionary relationships and paleo-distributional patterns of P. cayana in Mexico. Divergence time estimates suggest that the split between P. c. mexicana and P. c. thermophila occurred during the Pleistocene, about 1.24 Mya. Morphometrics of Mexican subspecies indicate that tail length and the white tail tips are significantly longer in P. c. mexicana, while P. c. thermophila has a longer and wider bill. Ecological niche analyses indicated that niche similarity between both lineages was lower than expected by chance, while low values on cross-prediction tests suggested that the two lineages have inhabited different environmental spaces since at least the Late Pleistocene. The ecological niche difference may also be associated with a steep humidity gradient, suggesting a significant contemporary environmental barrier. Overall, our results indicate that P. c. mexicana and P. c. thermophila have divergent evolutionary histories; therefore, the current taxonomic status of the Piaya populations in Mexico reflects neither their evolutionary relationships nor their apparent divergence. Our results suggest a major split in the polytypic P. cayana coinciding with the Andes, suggesting that the western endemic P. c. mexicana and P. c. thermophila are best treated as separate species-level taxa.
Peter Kovalik
Well-known member
Proposal (984) to SACC
Treat Piaya mexicana and P. “circe” as separate species from P. cayana (Squirrel Cuckoo)
Treat Piaya mexicana and P. “circe” as separate species from P. cayana (Squirrel Cuckoo)
Jim LeNomenclatoriste
Je suis un mignon petit Traquet rubicole
Why circe is in inverted commas?Proposal (984) to SACC
Treat Piaya mexicana and P. “circe” as separate species from P. cayana (Squirrel Cuckoo)
l_raty
laurent raty
Why circe is in inverted commas?
The split species would include circe and mehleri which were published simultaneously (Bonaparte 1850). The author suggests the precedence between these two names is not fixed, and uses "circe" provisionally.
Jim LeNomenclatoriste
Je suis un mignon petit Traquet rubicole
Historical contingency influences the diversity of feather nanostructures in cuckoos
Structural coloration is widespread in animals, yet we know relatively little about its evolution and development. While previous studies have explored adaptive functions of structural color, a key gap is our lack of understanding of how historical contingency (path-dependency of biological...
Abstract
Structural coloration is widespread in animals, yet we know relatively little about its evolution and development. While previous studies have explored adaptive functions of structural color, a key gap is our lack of understanding of how historical contingency (path-dependency of biological processes) influences the loss and gain of this complex trait. We shed light on this question by describing feather nanostructures responsible for plumage colors in the cuckoos (family Cuculidae), a group with widespread occurrence of shiny, metallic plumage (metallic luster). The melanosomes found in feathers with metallic luster have specialized shapes: hollow rods, thin solid rods, hollow platelets, or solid platelets. In contrast, it is generally assumed that drably colored feathers possess thick, rod-shaped melanosomes. However, we uncover that this assumption is unfounded in cuckoos. We describe metallic luster in the plumages of 126 cuckoo species and map its phylogenetic distribution. This reveals that metallic luster is widespread in cuckoos but has likely been lost several times. We then use transmission electron microscopy to describe the feather nanostructures of 21 cuckoo species. Surprisingly, the drab feathers of many cuckoo species contain melanosomes with specialized shapes. We propose that historical contingency greatly influences nanostructure diversity in cuckoos. Specialized melanosome shapes can be retained in the plumages of drab species, potentially making it easier for metallic luster to evolve again in the future. This discovery supports the idea that historical contingency plays a key role in shaping the evolution of plumage color diversity.
Acanthis
Well-known member
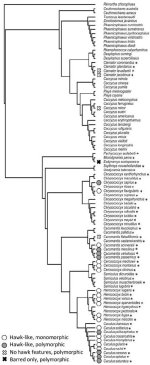
Would I be right in thinking that this paper has the first Cuculidae tree formed from genetic data?
Historical contingency influences the diversity of feather nanostructures in cuckoos
Structural coloration is widespread in animals, yet we know relatively little about its evolution and development. While previous studies have explored adaptive functions of structural color, a key gap is our lack of understanding of how historical contingency (path-dependency of biological...www.biorxiv.org
Abstract
Structural coloration is widespread in animals, yet we know relatively little about its evolution and development. While previous studies have explored adaptive functions of structural color, a key gap is our lack of understanding of how historical contingency (path-dependency of biological processes) influences the loss and gain of this complex trait. We shed light on this question by describing feather nanostructures responsible for plumage colors in the cuckoos (family Cuculidae), a group with widespread occurrence of shiny, metallic plumage (metallic luster). The melanosomes found in feathers with metallic luster have specialized shapes: hollow rods, thin solid rods, hollow platelets, or solid platelets. In contrast, it is generally assumed that drably colored feathers possess thick, rod-shaped melanosomes. However, we uncover that this assumption is unfounded in cuckoos. We describe metallic luster in the plumages of 126 cuckoo species and map its phylogenetic distribution. This reveals that metallic luster is widespread in cuckoos but has likely been lost several times. We then use transmission electron microscopy to describe the feather nanostructures of 21 cuckoo species. Surprisingly, the drab feathers of many cuckoo species contain melanosomes with specialized shapes. We propose that historical contingency greatly influences nanostructure diversity in cuckoos. Specialized melanosome shapes can be retained in the plumages of drab species, potentially making it easier for metallic luster to evolve again in the future. This discovery supports the idea that historical contingency plays a key role in shaping the evolution of plumage color diversity.
I don't recall any others
Jim LeNomenclatoriste
Je suis un mignon petit Traquet rubicole
There are others but I find the trees a bit strange in this articleWould I be right in thinking that this paper has the first Cuculidae tree formed from genetic data?
I don't recall any others
Acanthis
Well-known member
In what way?There are others but I find the trees a bit strange in this article
So many puzzling relationships within Cuculinae. It would be fun to delve into those.
A time-calibrated tree is high on my wish list for this group.
Last edited:
Jim LeNomenclatoriste
Je suis un mignon petit Traquet rubicole
You can compare this figure with that of the archived manuscriptIn what way?
So many puzzling relationships within Cuculinae. It would be fun to delve into those.
A time-calibrated tree is high on my wish list for this group.
Attachments
Users who are viewing this thread
Total: 2 (members: 0, guests: 2)