Fred Ruhe
Well-known member

Marco Pavia, 2020
Palaeoenvironmental reconstruction of the Cradle of Humankind during the Plio-Pleistocene transition, inferred from the analysis of fossil birds from Member 2 of the hominin-bearing site of Kromdraai (Gauteng, South Africa)
Quaternary Science Reviews. 248: Article 106532. doi:10.1016/j.quascirev.2020.106532
Highlights: https://www.sciencedirect.com/science/article/abs/pii/S0277379120304947?via=ihub
The fossil bird association of Kromdraai Member 2 is described in detail.
A great avian diversity (25 taxa, 2 new for the science) has been detected.
The high numbers of rock-dweller birds reinforce the presence of a cliff.
The presented results modify the palaeoenvironmental reconstruction of Kromdraai.
The importance of fossil birds as palaeoenvironmental proxies is confirmed.
Abstract
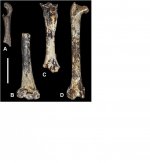
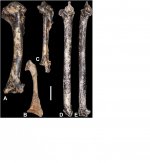
The results of the analysis of the bird assemblage from Member 2 of the Paranthropus robustus type-locality of Kromdraai are presented here, and the data are used to infer the environmental context of the Plio-Pleistocene transition in the Cradle of Humankind. The study of more than 800 remains revealed the presence of 25 bird taxa, 2 of them described as new to science. For some of the taxa, Kromdraai represents the oldest fossil record so far, if not the first ever. This relatively diverse bird association comprises terrestrial taxa dominated by rock-dwelling birds, such as Geronticus thackerayi and Tyto cf. alba, followed by open grassland birds, such as Francolinus sp. and Turnix sp., freshwater birds, ducks and waders, and forest birds, such as Accipiter melanoleucos and Treron sp.. The presence of these taxa, and their percentages in terms of the number of bones and individuals, point to the presence of a cliff or important rocky outcrop very close to the fossil site with the presence of a waterbody and a gallery forest. The grassland birds and their numbers confirm the presence of open grassland or savannah, which was already indicated by the analysis of large mammals. These results shed new light on the environment of the Cradle of Humankind during the Plio-Pleistocene transition and confirm the validity of the birds as palaeoenvironmental proxies.
Marco Pavia describes two new species: Glaucidium ireneae sp. n. and Corvus bragai sp. n.
Enjoy,
Fred
Palaeoenvironmental reconstruction of the Cradle of Humankind during the Plio-Pleistocene transition, inferred from the analysis of fossil birds from Member 2 of the hominin-bearing site of Kromdraai (Gauteng, South Africa)
Quaternary Science Reviews. 248: Article 106532. doi:10.1016/j.quascirev.2020.106532
Highlights: https://www.sciencedirect.com/science/article/abs/pii/S0277379120304947?via=ihub
The fossil bird association of Kromdraai Member 2 is described in detail.
A great avian diversity (25 taxa, 2 new for the science) has been detected.
The high numbers of rock-dweller birds reinforce the presence of a cliff.
The presented results modify the palaeoenvironmental reconstruction of Kromdraai.
The importance of fossil birds as palaeoenvironmental proxies is confirmed.
Abstract
The results of the analysis of the bird assemblage from Member 2 of the Paranthropus robustus type-locality of Kromdraai are presented here, and the data are used to infer the environmental context of the Plio-Pleistocene transition in the Cradle of Humankind. The study of more than 800 remains revealed the presence of 25 bird taxa, 2 of them described as new to science. For some of the taxa, Kromdraai represents the oldest fossil record so far, if not the first ever. This relatively diverse bird association comprises terrestrial taxa dominated by rock-dwelling birds, such as Geronticus thackerayi and Tyto cf. alba, followed by open grassland birds, such as Francolinus sp. and Turnix sp., freshwater birds, ducks and waders, and forest birds, such as Accipiter melanoleucos and Treron sp.. The presence of these taxa, and their percentages in terms of the number of bones and individuals, point to the presence of a cliff or important rocky outcrop very close to the fossil site with the presence of a waterbody and a gallery forest. The grassland birds and their numbers confirm the presence of open grassland or savannah, which was already indicated by the analysis of large mammals. These results shed new light on the environment of the Cradle of Humankind during the Plio-Pleistocene transition and confirm the validity of the birds as palaeoenvironmental proxies.
Marco Pavia describes two new species: Glaucidium ireneae sp. n. and Corvus bragai sp. n.
Enjoy,
Fred
Last edited: