Family Aegothelidae Bonaparte, 1853
Genus Aegotheles Vigors & Horsfield, 1827
Aegotheles zealandivetus sp. nov.
urn:lsid:zoobank.org:act:89951097-E888-4589-9FD2-8E928B40FAF0.
Holotype: NMNZ S.52917, a distal right tarsometatarsus, with part of the shaft and three trochleae exhibiting slight wear but otherwise well-preserved (Fig. 1A–E).
Etymology: From Zealandia, the continent on which New Zealand is emergent and vetus Latin adjective for old, ancient, or former, to reflect the fact that the new species was an old inhabitant of Zealandia, masculine.
Type locality: Bed HH1b, c.10 cm thick sand and coarse cobble layer, 9.5–9.58 m above base of Bannockburn Formation, Trench excavation, foot of hill 50 m across terrace from riverbank at 44.54.493°S; 169.85844°E; Manuherikia River, Home Hills Station, Otago, New Zealand. NZ Fossil Record File Number H41/f0103.
Stratigraphy and age: Bannockburn Formation, Manuherikia Group, early Miocene (19–16 Ma).
Diagnosis: The new taxon is an aegothelid having the following unique combination of characters for the distal tarsometatarsus: The fossa metatarsi I is deep, broad, not impacting on the medial profile of the shaft; the foramen vasculare distale is small, close to the incisura lateralis, and preceded by a short shallow sulcus; the trochleae diverge asymmetrically; the trochlea metatarsi II is directed somewhat medially making the medial profile concave; the trochlea metatarsi IV is slightly offset from the shaft, directed distally, resulting in convex lateral profile; the dorsal surface proximally adjacent to trochlea metatarsi III is flat, lacking a sulcus extending proximally from the incisura medialis; the distal projection of trochlea metatarsi II is slightly less than that of trochlea metatarsi IV, with 25% of the length of trochlea metatarsi III exceeding both; the trochlea metatarsi II lacks a groove dorsally; the trochlea metatarsi IV has a shallow groove distally but none dorsally; and the trochlea metatarsi III is deeply grooved dorsally, distally and plantarly, deeper than wide, with rims parallel in plantar aspect.
Differential diagnosis: Differs from Aegotheles savesi, A. novaezealandiae, A. salvadorii and A. cristatus on account of the following features: the dorsal surface proximal to trochlea metatarsi III is flat and merges with an evenly convex facies extending to the medial margin proximal to the trochlea metatarsi II without interruption by a groove extending from the intertrochlear incision (as opposed to being convex with a deep groove); the foramen vasculare distale is smaller; the trochlea metatarsi II is shorter than the trochlea metatarsi IV (as opposed to a relatively longer trochlea metatarsi II that distally exceeds trochlea metatarsi IV, or which may distally exceed trochlea metatarsi III in A. novaezealandiae).
Differs from A. novaezealandiae on account of: the trochlea metatarsi IV extends distally slightly past the trochlea metatarsi II; the trochlea metatarsi IV projects distally straight (not directed laterally); the fossa metatarsi I is larger and more excavated.
Differs from A. savesi on account of: the dorsal surface is flatter; the trochlea metatarsi II does not equal or surpass distally the trochlea metatarsi IV in medial/dorsal views; the trochlea metatarsi II is more plantarly retracted; the projection on the medial rim of the trochlea metatarsi II is smaller, even allowing for damage (A. savesi has a robust plantar flange); the trochlea metatarsi II is broader mediolaterally (dorsal view); the rims of the trochleae are more pronounced; there is a deeper furrow on trochlea metatarsi III, with rims that are parallel and not tapered in plantar aspect (A. savesi has proximally convergent rims); the trochlea metatarsi IV has a groove distally (lacking) and its outer rim lacks plantar projection (in A. savesi, the outer rim has a narrow flange extending about 30% of its depth plantar to the inner rim); the foramen vasculare distale is round, smaller and situated closer to incisura intertrochlearis; the fossa metatarsi I is wider/broader; and the cristae plantares medialis et lateralis is stronger.
Differs from Aegotheles cristatus and A. salvadorii on account of: larger size (Table 1); the trochlea metatarsi II is not as medially splayed; the trochlea metatarsi IV is directed mainly distally (more laterally) and is more laterally positioned relative to the shaft so that the profile is convex (versus straight in A. cristatus and A. salvadorii); the trochlea metatarsi III is deeply grooved; the trochlea metatarsi IV is shallowly grooved distally (in A. cristatus and A. salvadorii, all trochleae lack a groove both dorsally and distally).
Most similar to those of A. crinifrons and A. insignis, especially in the minimal development of a sulcus and ridge that extends proximally to the incisura medialis and the trochlea metatarsi III, respectively, and in the trochlea metatarsi II being comparatively short. Differs from A. crinifrons on account of: the shaft is more slender; the trochlea metatarsi II is shorter and level with the trochlea metatarsi IV; the trochleae metatarsorum III et IV are grooved; the trochlea metatarsi III has rims that are parallel in plantar view (convergent proximally). Differs from A. insignis on account of: the shaft is more slender; the trochlea metatarsi II is longer (IV much exceeds II in insignis); the trochlea metatarsi III is deeply grooved dorsally (lacking); the foramen vasculare distale is relatively larger.
Aegotheles tatei is the only similar-sized aegothelid that the specimen was not compared with, but it was considered conspecific with A. insignis until recently, so is probably very similar to that taxon; moreover, all these modern birds are separated from the fossil taxon by a minimum of 16 million years so are unlikely to be conspecific.
Referred specimens: All from beds on the East bank of the Manuherikia River, Home Hills Station, St Bathans, Otago, NZ. NMNZ S.42800, a proximal right tarsometatarsus, Bed HH1a, 6.88–7.0 m above base of Bannockburn Formation, at
44.907944S, 169.858222E. NZ Fossil Record File Number H41/f88. (Schwarzhans et al. 2012; Worthy et al. 2022). CM 2013.18.1127, cranial part of a right coracoid (Fig. 2A–C), Bed HH2, 21.02–21.48 m above the base of the Bannockburn Formation, at 44.907861°S 169.857389°E; NZ Fossil Record File Number H41/f87. NMNZ S.51974, proximal right ulna (Fig. 2G), Bed HH1b, details as per type locality. NMNZ S.52761, distal right tibiotarsus (Fig. 2J), Bed HH1b, details as per type locality.
Measurements (mm): Holotype, NMNZ S.52917, distal right tarsometatarsus, preserved length 12.0; shaft width 2.1; shaft depth 1.8; distal width 5.1. NMNZ S.42800, proximal right tarsometatarsus: preserved length 11.0; proximal width 4.3; proximal depth 4.1 mm; least shaft width 1.6 mm. NMNZ S.51974, proximal right ulna: maximum proximal width 4.5. CM 2013.18.1127, cranial part right coracoid: preserved length 12.6; cotyla scapularis to proximal side facies articularis humeralis 3.5. NMNZ S.52761, distal right tibiotarsus: distal width 4.4; proximodistal length condylus lateralis 3.1; proximodistal length condylus medialis 2.9; depth condylus medialis 4.1. See Table 1.
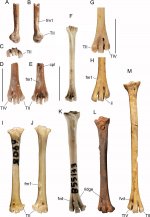
Fig. 1 Tarsometatarsi of species of Aegotheles. Aegotheles zealandivetus sp. nov. NMNZ S.52917, distal right tarsometatarsus (A–E) in lateral (A), medial (B), distal (C),
dorsal (D), plantar (E), aspects; Aegotheles cristatus SAM B.55133 (F, K); Aegotheles
novaezealandiae CM Av16996 (holotype), left tarsometatarsus (reversed) (H, M); A. novaezealandiae CM Av17503 (paratype), distal left tarsometatarsus (reversed) (G). A. insignis YPM ORN 104654 (I, J); A. savesi right tarsometatarsus, (IANCP unregistered: 100–110 cm Testpit 3, Pindai, New Caledonia) (L). Abbreviations: cpl,
crista plantaris lateralis; fm1, fossa metatarsi I; fvd, foramen vasculare distale; il, incisura intertrochlearis lateralis; TII, trochlea metatarsi II; TIV, rochlea metatarsi IV. Scale bars are 10 mm, and apply to images A–H. Images I–M are scaled so the distal width is the same as in (D, E)
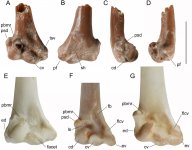
Fig. 2 Bones of species of Aegotheles: coracoids (A–D); ulnae (E–G); and tibiotarsi (H–J). Aegotheles zealandivetus sp. nov. cranial part right coracoid CM 2013.18.1127 (A–C), proximal right ulna NMNZ S.51974 (G), distal right tibiotarsus NMNZ S.52761 (J); Aegotheles cristatus SAM B.55133 (D, F, H), A. novaezealandiae CM Av28220 (paratype), proximal left ulna [reversed] (E), CM Av18240, distal left tibiotarsus reversed (K); A. insignis YPM ORN 104654, surface scan of specimen (I). Abreviations: cs, cotyla scapularis; cv, cotyla ventralis; dsf, distal sulcus of musculus fibularis; el, tuberositas retinaculi extensoris lateralis; em, tuberositas retinaculi extensoris medialis; fah, facies articularis humeralis; fs, foramen nervi supracoracoidei; ic, incisura intercondylaris; lc, condylus lateralis; mc, condylus nedialis; ol, olecranon; pcd, processus cotyla dorsalis; tlcv, tuberculum ligamentum collateralis ventralis; arrow points distal end of the processus procoracoideus. Scale
bars = 10 mm