Fred Ruhe
Well-known member

Ellen K. Mather, Michael S. V. Lee & Trevor H. Worthy, 2022
A new look at an old Australian raptor places “Taphaetus” lacertosus de Vis 1905 in the Old World vultures (Accipitridae: Aegypiinae)
Zootaxa: 1–23.
doi:10.11646/zootaxa.5168.1.1
Abstract and free pdf: A new look at an old Australian raptor places “ Taphaetus ” lacertosus de Vis 1905 in the Old World vultures (Accipitridae: Aegypiinae) | Zootaxa
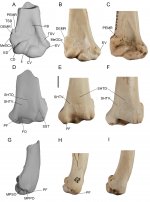
The Australian Pleistocene fossil record of the Accipitridae (hawks, eagles and Old World vultures) is sparse and poorly known. Only two extinct confirmed accipitrid species have been described for this time period; both have received little investigation since their description. One is “Taphaetus” lacertosus de Vis, 1905, described from a distal humerus and a quadrate from north-eastern South Australia. While this species was verified as an accipitrid in subsequent studies, its more precise taxonomic affinities have remained conjectural. In this study, a new analysis incorporating newly referred material and phylogenetic analyses using a wide range of accipitriforms reveals that the lectotype humerus of “T.” lacertosus is an Old World vulture in the subfamily Aegypiinae. The associated quadrate, one of two original syntypes from which de Vis named this species, is of an indeterminate species of ardeid. We erect the novel genus Cryptogyps, to accommodate the species ‘lacertosus’, as it cannot be placed in Taphaetus de Vis, 1891, because the type species of this genus, Uroaetus brachialis de Vis, 1889, was transferred back to the genus Uroaetus, a synonym of Aquila Brisson, by de Vis in 1905. Further, U. brachialis is now considered a synonym of Aquila audax (Latham, 1801). Moreover, Taphaetus de Vis, 1891 is a senior homonym of Taphaetus de Vis, 1905, type species Taphaetus lacertosus de Vis, 1905, making the 1905 version of the genus unavailable. Newly referred fossils from Wellington Caves (NSW) and the Nullarbor Plains (WA) reveal this taxon had a wide geographical range across Pleistocene Australia. The referred tarsometatarsus lacks hyper-developed trochleae, indicating that Cryptogyps lacertosus (de Vis, 1905) comb. nov., was probably a scavenger like other aegypiines. Identification of Cryptogyps lacertosus as an aegypiine significantly expands the palaeogeographical range of the Old World vultures, hitherto unknown in Australia. The avian guild of large, obligate scavenging birds of prey, is currently absent in the modern Australian biota, but its former presence is not surprising given the megafauna-rich communities of the Pleistocene.
Enjoy,
Fred
A new look at an old Australian raptor places “Taphaetus” lacertosus de Vis 1905 in the Old World vultures (Accipitridae: Aegypiinae)
Zootaxa: 1–23.
doi:10.11646/zootaxa.5168.1.1
Abstract and free pdf: A new look at an old Australian raptor places “ Taphaetus ” lacertosus de Vis 1905 in the Old World vultures (Accipitridae: Aegypiinae) | Zootaxa
The Australian Pleistocene fossil record of the Accipitridae (hawks, eagles and Old World vultures) is sparse and poorly known. Only two extinct confirmed accipitrid species have been described for this time period; both have received little investigation since their description. One is “Taphaetus” lacertosus de Vis, 1905, described from a distal humerus and a quadrate from north-eastern South Australia. While this species was verified as an accipitrid in subsequent studies, its more precise taxonomic affinities have remained conjectural. In this study, a new analysis incorporating newly referred material and phylogenetic analyses using a wide range of accipitriforms reveals that the lectotype humerus of “T.” lacertosus is an Old World vulture in the subfamily Aegypiinae. The associated quadrate, one of two original syntypes from which de Vis named this species, is of an indeterminate species of ardeid. We erect the novel genus Cryptogyps, to accommodate the species ‘lacertosus’, as it cannot be placed in Taphaetus de Vis, 1891, because the type species of this genus, Uroaetus brachialis de Vis, 1889, was transferred back to the genus Uroaetus, a synonym of Aquila Brisson, by de Vis in 1905. Further, U. brachialis is now considered a synonym of Aquila audax (Latham, 1801). Moreover, Taphaetus de Vis, 1891 is a senior homonym of Taphaetus de Vis, 1905, type species Taphaetus lacertosus de Vis, 1905, making the 1905 version of the genus unavailable. Newly referred fossils from Wellington Caves (NSW) and the Nullarbor Plains (WA) reveal this taxon had a wide geographical range across Pleistocene Australia. The referred tarsometatarsus lacks hyper-developed trochleae, indicating that Cryptogyps lacertosus (de Vis, 1905) comb. nov., was probably a scavenger like other aegypiines. Identification of Cryptogyps lacertosus as an aegypiine significantly expands the palaeogeographical range of the Old World vultures, hitherto unknown in Australia. The avian guild of large, obligate scavenging birds of prey, is currently absent in the modern Australian biota, but its former presence is not surprising given the megafauna-rich communities of the Pleistocene.
Enjoy,
Fred